
El cerebelo representa un papel fundamental en la coordinación temporal de las actividades motoras y en el paso suave y rápido desde un movimiento muscular al siguiente. También sirve para regular la intensidad de la contracción muscular cuando varía la carga a la que se encuentra sometida, y controla las interacciones instantáneas que son necesarias entre los grupos musculares agonistas y antagonistas.
Los ganglios basales ayudan a planificar y controlar los patrones complejos de movimiento muscular. Regulan las intensidades relativas de cada movimiento independiente, su dirección y la ordenación de los movimientos paralelos y sucesivos múltiples destinados a alcanzar un objetivo
motor específico complicado.
EL CEREBELO Y SUS FUNCIONES MOTORAS
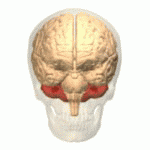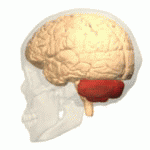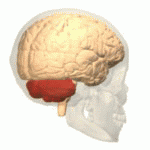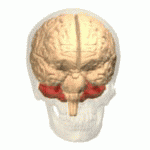
El cerebelo resulta especialmente vital durante las actividades musculares rápidas como correr, escribir a máquina, tocar el piano e incluso conversar. La desaparición de este componente del encéfalo puede provocar una falta de coordinación casi total de estas tareas aun cuando su pérdida no ocasione la parálisis de ningún músculo.

El cerebelo recibe constantemente información actualizada acerca de la secuencia deseada de contracciones musculares desde las áreas encefálicas de control motor; también le llega una información sensitiva continua desde las porciones periféricas del organismo, que comunica las variaciones sucesivas en el estado de cada una de ellas: su posición, la velocidad de movimiento, las fuerzas que actúan sobre ella, etc. A continuación, el cerebelo contrasta los movimientos reales descritos por la información sensitiva periférica de retroalimentación con los movimientos pretendidos por el sistema motor. Si la comparación entre ambos no resulta satisfactoria, entonces devuelve unas señales subconscientes instantáneas de corrección hacia el sistema motor para aumentar o disminuir los niveles de activación de cada músculo específico.
LA UNIDAD FUNCIONAL DE LA CORTEZA CEREBELOSA: LA CÉLULA DE PURKINJE Y LA CÉLULA NUCLEAR PROFUNDA

La salida desde el cerebelo tiene lugar a través de una célula nuclear profunda. Esta célula está sometida permanentemente a unas influencias excitadoras e inhibidoras. Las influencias excitadoras emanan de sus conexiones directas con fibras aferentes que llegan al cerebelo desde el
encéfalo o desde la periferia; la inhibidora procede en su totalidad de la célula de Purkinje situada en la corteza cerebelosa.
Las fibras trepadoras nacen en su integridad en las olivas inferiores del bulbo raquídeo. Hay una fibra trepadora por cada 5 a 10 células de Purkinje. Después de enviar ramas hacia varias células nucleares profundas, estas fibras siguen su camino hacia las capas superficiales de la corteza cerebelosa, donde realizan unas 300 sinapsis con los somas y las dendritas de cada célula de Purkinje.
La fibra trepadora se distingue por el hecho de que un solo impulso suyo siempre generará un solo tipo peculiar de potencial de acción prolongado (hasta 1 s) en cada célula de Purkinje con la que conecta, cuya configuración comienza con una descarga potente y va seguida de un reguero de descargas secundarias cada vez más débiles. Este potencial de acción se llama descarga compleja.
Además de las células nucleares profundas, las células de los granos y las células de Purkinje, en el cerebelo hay otros dos tipos de neuronas: las células en cesta y las células estrelladas, que son células inhibidoras con axones cortos. Ambas están situadas en la capa molecular de la corteza cerebelosa, ubicadas entre las pequeñas fibras paralelas y estimuladas por ellas. Estas células a su vez envían unos axones perpendiculares a dichas fibras paralelas que ocasionan una inhibición lateral de las células de Purkinje adyacentes, lo que afina la señal del mismo modo que el mecanismo de inhibición lateral acentúa el contraste de las señales en otros muchos circuitos neuronales del sistema nervioso.
SEÑALES DE ENCENDIDO – APAGADO Y APAGADO – ENCENDIDO EMITIDAS POR EL CEREBELO

La función típica del cerebelo consiste en contribuir a suministrar unas señales rápidas de encendido para los músculos agonistas y simultáneamente unas señales recíprocas de apagado para los antagonistas al comenzar un movimiento. A continuación, cuando se acerca su final, el cerebelo es básicamente el responsable de sincronizar y ejecutar las señales de apagado dirigidas a los agonistas y de encendido para los antagonistas.
Vamos a suponer que el patrón de encendido-apagado en la contracción de los agonistasantagonistas al comienzo del movimiento comienza con las señales procedentes de la corteza cerebral. Estas señales recorren vías no cerebelosas en el tronco del encéfalo y la médula que llegan directamente hasta el músculo agonista para poner en marcha la contracción inicial.
Al mismo tiempo, unas señales paralelas acceden al cerebelo por medio de las fibras musgosas pontinas. Una rama de cada fibra musgosa va directamente hasta las células nucleares profundassituadas en el núcleo dentado o en otros núcleos cerebelosos profundos, con lo que se devuelve al instante una señal excitadora hacia el sistema motor corticoespinal, ya sea mediante los impulsos de regreso hasta la corteza cerebral a través del tálamo o recurriendo al circuito neuronal en el tronco del encéfalo, con objeto de respaldar la señal de contracción muscular que ya se había puesto en
marcha en la corteza cerebral. Como consecuencia de ello, pasados unos pocos milisegundos, la señal de encendido adquiere aún mayor potencia que la que tenía al comienzo debido a que es el resultado de sumar las señales corticales más las cerebelosas. Este es el efecto normal cuando el cerebelo se encuentra íntegro, pero en su ausencia desaparece la señal de refuerzo adicional secundaria. Esta contribución cerebelosa vuelve la contracción muscular de encendido mucho más enérgica de lo que sería si su participación no existiera.
Es preciso tener presente que el cerebelo contiene varios tipos más de células inhibidoras aparte de las células de Purkinje. Aún quedan por determinar las funciones de algunas de ellas; por añadidura, también podrían ocupar algún lugar en la inhibición lateral de los músculos antagonistas al comienzo de un movimiento y en su posterior excitación cuando acabe su realización.
ESPINOCEREBELO: CONTROL POR RETROALIMENTACION DE LOS MOVIMIENTOS DISTALES DE LAS EXTREMIDADES A TRAVÉS DE LA CORTEZA CEREBELOSA INTERMEDIA Y EL NUCLEO INTERPUESTO.

Cuando se realiza un movimiento la zona intermedia de cada hemisferio cerebeloso recibe dos tipos de datos: 1) información procedente de la corteza
cerebral motora y del núcleo rojo mesencefálico, que avisa al cerebelo sobre el plan de movimiento secuencial pretendido durante las fracciones de segundo siguientes, y 2) información de retroalimentación procedente de las porciones periféricas del cuerpo, en especial de los propioceptores distales de las extremidades, que transmite al cerebelo los movimientos reales resultantes.
Una vez que la zona intermedia del cerebelo ha comparado los movimientos deseados con los movimientos reales, las células nucleares profundas del núcleo interpuesto envían unas señales eferentes correctoras: 1) de vuelta hacia la corteza cerebral motora a través de los núcleos de relevo en el tálamo, y 2) hacia la porción magnocelular (inferior) del núcleo rojo que da origen al fascículo rubroespinal. Este último, a su vez, se suma al fascículo corticoespinal en su inervación de las motoneuronas más laterales contenidas en las astas anteriores de la sustancia gris de la médula espinal, las células que controlan las partes distales de las extremidades, en especial las manos y los dedos.
CEREBROCEREBELO: FNUNCIÓN DE LA GRAN ZONA LATERAL DEL HEMISFERIO CEREBELOSO PARA PLANIFICAR, ORDENAR Y SINCRONIZAR LOS MOVIMIENTOS COMPLEJOS

En el ser humano, las zonas laterales de los dos hemisferios cerebelosos están muy desarrolladas e hipertrofiadas. Esta característica encaja con las capacidades del hombre para planificar y ejecutar patrones secuenciales complicados de movimiento, especialmente con las manos y con los dedos, y
para hablar. Con todo, estas grandes zonas laterales de los hemisferios cerebelosos carecen de una vía de entrada directa para la información procedente de las porciones periféricas del cuerpo. Asimismo, casi toda la comunicación entablada entre dichas áreas cerebelosas laterales y la corteza cerebral no se dirige a la corteza motora primaria sino al área premotora y las áreas somatosensitivas primaria y de asociación.
La planificación de los movimientos secuenciales exige que las zonas laterales de los hemisferios estén en contacto con las porciones sensitivas y premotoras de la corteza cerebral, y esto requiere una comunicación bidireccional entre estas áreas corticales cerebrales y las regiones correspondientes de los ganglios basales. Parece que el «plan» de los movimientos secuenciales en realidad comienza en las áreas sensitivas y premotoras de la corteza cerebral, y desde allí se transmite hacia las zonas laterales de los hemisferios cerebelosos. Entonces, en medio de un gran tráfico de doble sentido entre el cerebelo y la corteza cerebral, las señales motoras oportunas proporcionan la transición entre una secuencia de movimientos y la siguiente.
GANGLIOS BASALES Y SUS FUNCIONES MOTORAS

Los ganglios basales, igual que el cerebelo, constituyen otro sistema motor auxiliar que en general no funciona por su cuenta sino íntimamente vinculado con la corteza cerebral y el sistema de control motor corticoespinal. De hecho, reciben la mayoría de sus señales aferentes desde la corteza cerebral y también devuelven casi todas sus señales eferentes a esta estructura.
A cada lado del encéfalo, están formados por el núcleo caudado, el putamen, el globo
pálido, la sustancia negra y el núcleo subtalámico. Se encuentran situados básicamente en una posición lateral y alrededor del tálamo, ocupando una gran parte de las regiones internas de ambos hemisferios cerebrales. Casi todas las fibras nerviosas sensitivas y motoras que conectan la corteza cerebral con la médula espinal atraviesan el área que queda entre los elementos más voluminosos de los ganglios basales, el núcleo caudado y el putamen. Este espacio se llama cápsula interna del cerebro. Tiene importancia en lo que atañe a nuestra explicación actual debido a la intensa asociación que existe entre los ganglios basales y el sistema corticoespinal para el ejercicio del control motor.
Uno de los principales cometidos que cumplen los ganglios basales en el control motor consiste en su funcionamiento vinculado al sistema corticoespinal con objeto de controlar los patrones complejos de la actividad motora.
Algunos patrones que requieren el funcionamiento de los ganglios basales son los encargados de efectuar las siguientes actividades: cortar un papel con unas tijeras, fijar un clavo a martillazos, meter un balón de baloncesto en la canasta, dar un pase de fútbol, lanzar una pelota de béisbol, quitar tierra con una pala, la mayoría de las diversas facetas de la vocalización, los movimientos controlados de los ojos y prácticamente cualquier otra de las acciones que exijan una cierta destreza, la mayoría de ellas ejecutadas de forma subconsciente.
El término conocimiento o cognición se refiere a los procesos de pensamiento del encéfalo, que emplean las señales sensitivas llegadas al cerebro más la información ya almacenada en la memoria. La mayoría de nuestras acciones motoras se dan como consecuencia de los pensamientos generados en la mente, fenómeno llamado control cognitivo de la actividad motora. El núcleo caudado representa un papel fundamental en este proceso.
