
Control local del flujo sanguíneo en respuesta a las necesidades tisulares

Un principio fundamental de la función circulatoria es que la mayoría de los tejidos tienen la capacidad de controlar su propio flujo sanguíneo local en proporción a sus necesidades metabólicas concretas. Algunas de las necesidades específicas de flujo sanguíneo en los tejidos incluyen aspectos como:
- Aporte de oxígeno a los tejidos.
- Aporte de otros nutrientes, como glucosa, aminoácidos y ácidos grasos.
- Eliminación de dióxido de carbono de los tejidos.
- Eliminación de iones hidrógeno de los tejidos.
- Mantenimiento de las concentraciones adecuadas de iones en los tejidos.
- Transporte de varias hormonas y otras sustancias a los distintos tejidos.
Algunos órganos tienen necesidades especiales. Por ejemplo, el flujo sanguíneo de la piel determina la pérdida de calor corporal y, de esta forma, se controla la temperatura. Además, el aporte de cantidades adecuadas de plasma sanguíneo a los riñones permite que estos filtren y excreten los productos de desecho del organismo y regulen los volúmenes de líquidos y electrólitos.
Importancia del control del flujo sanguíneo por los tejidos locales
Puede plantearse una pregunta sencilla: ¿por qué no se proporciona un flujo sanguíneo muy importante a cada tejido del organismo, que sea siempre suficiente para cubrir las necesidades de los tejidos, con independencia de que la actividad tisular sea pequeña o grande? La respuesta es igualmente sencilla: dicho mecanismo requeriría muchas más veces el flujo sanguíneo que el corazón puede bombear.
En estudios experimentales se ha demostrado que el flujo sanguíneo que llega a un tejido está regulado por la concentración mínima que cubrirá las necesidades tisulares, ni más, ni menos. Por ejemplo, en los tejidos en los que la necesidad más importante es la administración de oxígeno, el flujo sanguíneo siempre está controlado a un nivel que solo es ligeramente mayor de lo necesario para mantener la oxigenación tisular plena, pero nada más. Al controlar el flujo sanguíneo local de una forma tan exacta, los tejidos casi nunca padecen una deficiencia nutricional de oxígeno y, a pesar de ello, la carga de trabajo del corazón se mantiene al mínimo.
Mecanismos de control del flujo sanguíneo
El control del flujo sanguíneo local se puede dividir en dos fases: 1) control a
corto plazo, y 2) control a largo plazo. El control a corto plazo se consigue con
cambios rápidos de la vasodilatación o vasoconstricción local de las arteriolas,
metaarteriolas y esfínteres precapilares, que se producen en segundos o minutos para proporcionar con gran rapidez el mantenimiento del flujo sanguíneo tisular local apropiado.
El control a largo plazo significa cambios controlados lentos del flujo en un período de días, semanas o incluso meses. En general, estos cambios a largo plazo proporcionan un control aún mejor del flujo en proporción a las necesidades de los tejidos. Estos cambios se producen como consecuencia del incremento o descenso del tamaño físico y del número de vasos sanguíneos que nutren los tejidos.
Control a corto plazo del flujo sanguíneo local
La disponibilidad reducida de oxígeno incrementa el flujo sanguíneo tisular
Uno de los nutrientes metabólicos más necesarios de los tejidos es el oxígeno. El flujo sanguíneo tisular aumenta mucho siempre que disminuye la disponibilidad de oxígeno en los tejidos, por ejemplo: 1) a una gran altitud, en la cima de una montaña alta; 2) en caso de neumonía; 3) en el envenenamiento por monóxido de carbono (que deteriora la capacidad de la hemoglobina de transportar el oxígeno), o 4) en el envenenamiento por cianuro (que deteriora la capacidad del tejido de usar oxígeno).
El mecanismo por el cual los cambios en el metabolismo tisular o en la disponibilidad de oxígeno modifican el flujo sanguíneo en los tejidos no se comprende totalmente, si bien se han propuesto dos teorías principales: la teoría vasodilatadora y la teoría de la falta de oxígeno.
Teoría de la demanda de oxígeno para el control del flujo sanguíneo local
Aunque la teoría vasodilatadora es la más aceptada, hay varios hechos críticos que fundamentan otra teoría más expuesta por otros fisiólogos, que podríamos denominar la teoría de la demanda de oxígeno o, más exactamente, la teoría de la demanda de nutrientes (porque están implicados otros nutrientes, además del oxígeno). El oxígeno es uno de los nutrientes metabólicos necesarios para provocar la contracción muscular (con la necesidad también de otros nutrientes). Por tanto, es razonable creer que los vasos sanguíneos simplemente se relajarían en ausencia de una cantidad adecuada de oxígeno, dilatándose de forma natural. Además, el aumento de la utilización de oxígeno en los tejidos como consecuencia del aumento delmetabolismo podría, en teoría, disminuir la disponibilidad de oxígeno hacia las fibras musculares lisas de los vasos sanguíneos locales, lo cual también provocaría la vasodilatación local.
Autorregulación del flujo sanguíneo durante los cambios en la presión arterial: mecanismos «metabólicos» y «miógenos»
En algunos tejidos, como el encéfalo y el corazón, esta autorregulación es todavíacmás precisa. Durante casi un siglo se han mantenido dos opiniones que explicarían el mecanismo de autorregulación a corto plazo, la teoría metabólica y la teoría miógena. La teoría metabólica se puede entenderfácilmente si se aplican los principios básicos de regulación del flujo sanguíneo local que hemos comentado en las secciones previas. Es decir, cuando la presión arterial es demasiado elevada, el exceso de líquido proporciona demasiado oxígeno y demasiados nutrientes de otro tipo hacia los tejidos y
«lava» los vasodilatadores liberados por los tejidos. Estos nutrientes (en especial, el oxígeno), junto con el descenso en los niveles tisulares de vasodilatadores, provocan entonces la constricción de los vasos sanguíneos y el retorno del flujo casi a la normalidad, a pesar de que aumente la presión. No obstante, según esta teoría miógena habría otro mecanismo no relacionado con el metabolismo tisular que explicaría el fenómeno de la autorregulación. Esta teoría se basa en la observación de que el estiramiento brusco de los vasos sanguíneos pequeños provoca la contracción del músculo liso de la pared vascular durante unos segundos. Por tanto, se ha propuesto que cuando una presión arterial elevada estira el vaso se provoca, a su vez, una constricción vascular reactiva que reduce el flujo sanguíneo casi a la normalidad. Por el contrario, con presiones bajas el grado de estiramiento del vaso es menor, por lo que el músculo liso se relaja, reduce la resistencia vascular y ayuda a recuperar la normalidad del flujo.
La respuesta miógena es inherente al músculo liso vascular y puede producirse en ausencia de influencias nerviosas u hormonales. Es más pronunciada en las arteriolas, pero se puede ver también en arterias, vénulas, venas e incluso en vasos linfáticos. La contracción miógena se inicia por la despolarización vascular inducida por el estiramiento, que tiende a aumentar rápidamente la entrada de ion calcio desde el líquido extracelular hacia las células, provocando su contracción. Los cambios de la presión vascular también pueden abrir o cerrar otros canales iónicos que influyen en la contracción vascular. Se desconocen los mecanismos por los cuales los cambios de la
presión provocan la apertura o el cierre de los canales iónicos vasculares, pero es probable que consistan en efectos mecánicos de la presión sobre las proteínas extracelulares que están ancladas en los elementos del citoesqueleto de la pared vascular o en los propios canales iónicos. El mecanismo miógeno parece ser importante para prevenir el estiramiento excesivo de los vasos sanguíneos cuando aumenta la presión sanguínea. No obstante, también se desconoce la importancia de este mecanismo en la regulación del flujo sanguíneo porque este mecanismo de detección de la presión no puede detectar directamente los cambios del flujo sanguíneo en el tejido. En realidad, los factores metabólicos anulan el mecanismo miógeno cuando las demandas metabólicas de los tejidos están significativamente aumentados, como durante el ejercicio muscular enérgico, lo que provoca un incremento espectacular del flujo sanguíneo en el músculo esquelético.
Mecanismos especiales del control a corto plazo del flujo sanguíneo en
tejidos específicos
Aunque los mecanismos generales de control del flujo sanguíneo local que hemos comentado hasta ahora actúan en la mayoría de los tejidos del organismo, en algunos territorios especiales actúan otros mecanismos totalmente diferentes. Todos los mecanismos se comentarán en este texto en relación con cada órgano específico, pero hay dos que merecen una atención especial:

- En los riñones, el control del flujo sanguíneo se basa en gran medida en un mecanismo denominado retroalimentación tubuloglomerular, en el que una estructura epitelial del túbulo distal, la mácula densa, detecta la composición del líquido al inicio de dicho túbulo. Esta estructura se sitúa en la zona en que el túbulo distal se encuentra cerca de las arteriolas aferente y eferente del aparato yuxtaglomerular de la nefrona. Cuando se filtra demasiado líquido de la sangre a través del glomérulo hacia el sistema tubular, las señales de retroalimentación de la mácula densa provocan constricción de las arteriolas aferentes, reduciendo de esta forma tanto el flujo sanguíneo renal como la tasa de filtración glomerular a valores casi normales.
- En el cerebro, además del control del flujo sanguíneo dependiente de la concentración de oxígeno tisular, las concentraciones de dióxido de carbono y de iones hidrógeno tienen una gran importancia. El aumento de cualquiera de ellos dilata los vasos cerebrales y permite el lavado rápido del exceso de dióxido de carbono o de iones hidrógeno de los tejidos cerebrales. Este mecanismo es importante porque el nivel de excitabilidad del propio cerebro depende en gran medida del control exacto de las concentraciones de dióxido de carbono y del ion hidrógeno.

- En la piel, el control del flujo sanguíneo está relacionado estrechamente con la regulación de la temperatura corporal. El flujo cutáneo y subcutáneo regula la pérdida de calor del cuerpo mediante la determinación del flujo de calor desde el centro a la superficie del organismo, donde se pierde calor hacia el medio exterior. El flujo sanguíneo en la piel está controlado en gran medida por el sistema nervioso central a través de los nervios simpáticos.
Control del flujo sanguíneo tisular por medio de factores de relajación y
contracción de origen endotelial
Las células endoteliales que recubren los vasos sanguíneos sintetizan varias sustancias que, cuando se liberan, afectan al grado de relajación o contracción de la pared arterial. Para muchos de estos factores de relajación o contracción de origen endotelial, las funciones fisiológicas apenas se están empezando a comprender y, en la mayoría de los casos, todavía no se han desarrollado aplicaciones clínicas.
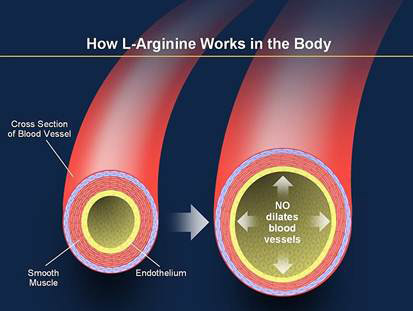
Óxido nítrico: un vasodilatador liberado por células endoteliales sanas
El más importante de los factores de relajación de origen endotelial es el óxido nítrico (NO), un gas lipófilo que es liberado por las células endoteliales como respuesta a diversos estímulos químicos y físicos. Las enzimas óxido nítrico sintasa de origen endotelial (eNOS) sintetizan el NO a partir de arginina y oxígeno y por reducción de nitrato inorgánico. Después de la difusión fuera de la célulaendotelial, el NO tiene una semivida en sangre de solo 6 s, aproximadamente, y actúa principalmente en los tejidos locales en los que es liberado.
El flujo de sangre a través de las arterias y arteriolas provoca fuerzas de cizallamiento sobre las células endoteliales por el arrastre viscoso de la sangre contra las paredes vasculares. NO relaja entonces los vasos sanguíneos, lo que es una ventaja porque los mecanismos metabólicos locales para el control del flujo sanguíneo tisular dilatan principalmente las arterias y arteriolas muy pequeñas en cada tejido.
Regulación a largo plazo del flujo sanguíneo
El flujo sanguíneo se ajusta solo en las tres cuartas partes de las necesidades adicionales de los tejidos, incluso después de la activación completa de estos mecanismos agudos. Por ejemplo, el flujo sanguíneo aumenta casi instantáneamente en un 100% cuando la presión arterial aumenta bruscamente desde 100 a 150 mmHg. En los 30 s a 2 min siguientes el flujo vuelve a disminuir hasta un 10-15% por encima del valor de control original.
Regulación del flujo sanguíneo por cambios en la «vascularización tisular»
Un mecanismo clave para la regulación del flujo sanguíneo local a largo plazo consiste principalmente en cambiar la cantidad de vascularización de los tejidos. Por ejemplo, la vascularización aumenta si el metabolismo de un tejido dado aumenta durante un período prolongado, en un proceso denominado generalmente angiogenia; si el metabolismo disminuye, la vascularización también lo hace. Se produce una reconstrucción física real de la vasculatura tisular para cubrir las necesidades de los tejidos. Esta reconstrucción es rápida (en días) en los animales muy jóvenes y también en un tejido de nuevo crecimiento, como en el tejido cicatricial o el tejido canceroso, pero es mucho más lenta en los tejidos antiguos y bien establecidos.
Función del oxígeno en la regulación a largo plazo

El oxígeno es importante no solo para el control a corto plazo del flujo sanguíneo local, sino también para el control a largo plazo. Un ejemplo es el aumento de la vascularización de los tejidos en los animales que viven en altitudes elevadas, donde el oxígeno atmosférico es bajo. En los recién nacidos prematuros que son tratados en tiendas de oxígeno con fines terapéuticos, el exceso de oxígeno provoca la interrupción casi inmediata del crecimiento vascular nuevo en la retina e incluso la degeneración de algunos de los vasos pequeños que ya se han formado. Cuando el niño es sacado de la tienda de oxígeno se produce un sobrecrecimiento explosivo de los vasos nuevos para
compensar el descenso brusco del oxígeno disponible. En realidad, el sobrecrecimiento es tal que los vasos retinianos sobrepasan la retina hacia el humor vítreo del ojo, lo que terminará por provocar ceguera (afección que se conoce con el nombre de fibroplasia retrolental).
Sustancias vasoconstrictoras
Noradrenalina y adrenalina

La noradrenalina es una hormona vasoconstrictora especialmente potente; la adrenalina es menos potente y en algunos tejidos provoca incluso una vasodilatación leve. (Un ejemplo especial de vasodilatación provocada por la adrenalina es el que tiene lugar para la dilatación coronaria durante el aumento de la actividad cardíaca.)
Cuando se estimula el sistema nervioso simpático en el cuerpo durante el estrés o el ejercicio, las terminaciones nerviosas simpáticas de cada tejido liberan noradrenalina, que excita al corazón y contrae las venas y las arteriolas.
Además, los nervios simpáticos de la médula suprarrenal provocan la secreción de noradrenalina y adrenalina en la sangre. Estas hormonas circulan entonces por todo el cuerpo y provocan casi los mismos efectos en la circulación que la estimulación simpática directa, con lo que se consigue un sistema de control doble: 1) estimulación nerviosa directa, y 2) efectos indirectos de la} noradrenalina y/o de la adrenalina en la sangre circulante.
Angiotensina II

La angiotensina II es otra sustancia vasoconstrictora potente. Tan solo una millonésima de gramo puede aumentar la presión arterial de un ser humano en 50 mmHg o más.
El efecto de angiotensina II contrae potentemente las pequeñas arteriolas. Si esta contracción sucede en un tejido aislado, el flujo sanguíneo de esa zona disminuirá mucho, aunque la importancia real de la angiotensina II es que normalmente actúa sobre muchas de las arteriolas del organismo al mismo tiempo, para aumentar la resistencia periférica total y reducir la excreción de sodio y agua en los riñones, lo que aumenta la presión arterial. Es decir, esta hormona tiene un papel fundamental en la regulación de la presión arterial.
Vasopresina

La vasopresina, que también se conoce como hormona antidiurética, es aún más potente que la angiotensina II como vasoconstrictora, por lo que se convierte en una de las sustancias constrictoras más potentes del organismo. Se forma en las células nerviosas del hipotálamo pero después es transportada distalmente a través de los axones nerviosos hacia la neurohipófisis, donde es finalmente segregada a la sangre.
Es evidente que la vasopresina podría tener efectos muy importantes sobre la función circulatoria. Sin embargo, dado que en la mayoría de las condiciones fisiológicas solo se segregan cantidades mínimas de vasopresina, se ha demostrado que la concentración de vasopresina en sangre circulante puede aumentar después de una hemorragia intensa, lo suficiente como para elevar la presión arterial hasta en 60 mmHg. En muchos casos, esta acción puede elevar por sí sola la presión arterial hasta la normalidad. La vasopresina tiene una función importante de aumentar la reabsorción de agua de los túbulos renales hacia la sangre (como se comenta en el capítulo 29) y ayuda, por tanto, a controlar el volumen de líquido corporal. De ahí viene el nombre de hormona antidiurética.
Sustancias vasodilatadoras
Bradicinina
Hay un grupo de sustancias denominadas cininas que provocan una vasodilatación potente cuando se forman en la sangre y en los líquidos tisulares de algunos órganos. Las cininas son pequeños polipéptidos que se escinden por enzimas proteolíticas a partir de α2- globulinas del plasma o los líquidos tisulares. Una enzima proteolítica de particular importancia para tal fin es la calicreína, que se encuentra en la sangre y los líquidos tisulares en una forma inactiva. Esta calicreína inactiva se activa por la maceración de la sangre, por la inflamación tisular o por otros efectos químicos o físicos similares. A medida que se va activando la calicreína actúa inmediatamente sobre la α2-globulina para liberar una cinina llamada calidina, que después se convierte en bradicinina gracias a las enzimas tisulares. Una vez formada, la bradicinina persiste durante solo unos minutos, porque se inactiva por la enzima carboxipeptidasa o por la enzima convertidora, la misma que participa en la activación de la angiotensina. La enzima calicreína activada se destruye por un inhibidor de la calicreína que también está presente en los líquidos corporales. La bradicinina provoca una dilatación arteriolar potente y aumenta la permeabilidad capilar. Por ejemplo, la inyección de 1 μm de bradicinina en la arteria braquial de una persona aumenta el flujo sanguíneo a través del brazo hasta en seis veces, e incluso cantidades menores inyectadas localmente en los tejidos pueden provocar un edema local importante como consecuencia del aumento de tamaño de los poros capilares.
Histamina

La histamina se libera esencialmente en todos los tejidos del organismo cuando sufren daños o se inflaman, o cuando se sufre una reacción alérgica. La mayor parte de la histamina deriva de los mastocitos en los tejidos dañados y de los basófilos en sangre. La histamina tiene un efecto vasodilatador potente sobre las arteriolas y, como la bradicinina, puede aumentar en gran medida la porosidad capilar permitiendo la pérdida tanto de líquidos como de proteínas plasmáticas hacia los tejidos. En muchas situaciones patológicas la dilatación arteriolar intensa y el aumento de la porosidad capilar producida por la histamina provoca la pérdida de cantidades enormes de líquido desde la circulación hacia los tejidos, induciendo el edema. Los efectos locales vasodilatadores y productores de edema de la histamina son especialmente prominentes durante las reacciones alérgicas.
