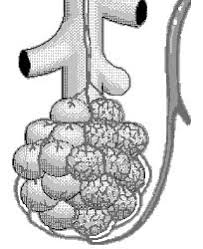
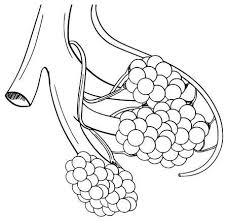
El pulmón tiene dos circulaciones: una circulación de bajo flujo y alta presión y una circulación de alto flujo y baja presión.
La circulación de bajo flujo y alta presión aporta la sangre arterial sistémica a la tráquea, el árbol bronquial incluidos los bronquíolos terminales, los tejidos de sostén del pulmón y las capas exteriores (adventicias) de las arterias y venas pulmonares. Las arterias bronquiales, que son ramas de la aorta torácica, irrigan la mayoría de esta sangre arterial sistémica a una presión solo ligeramente inferior a la presión aórtica.
La circulación de alto flujo y baja presión que suministra la sangre venosa de todas las partes del organismo a los capilares alveolares en los que se añade el oxígeno (O2) y se extrae el dióxido de carbono (CO2). La arteria pulmonar, que recibe sangre del ventrículo derecho, y sus ramas arteriales transportan sangre a los capilares alveolares para el intercambio gaseoso y a las venas pulmonares y después devuelven la sangre a la aurícula izquierda para su bombeo por el ventrículo izquierdo a través de la circulación sistémica.
ANATOMÍA FISIOLÓGICA DEL SISTEMA CIRCULATORIO PULMONAR
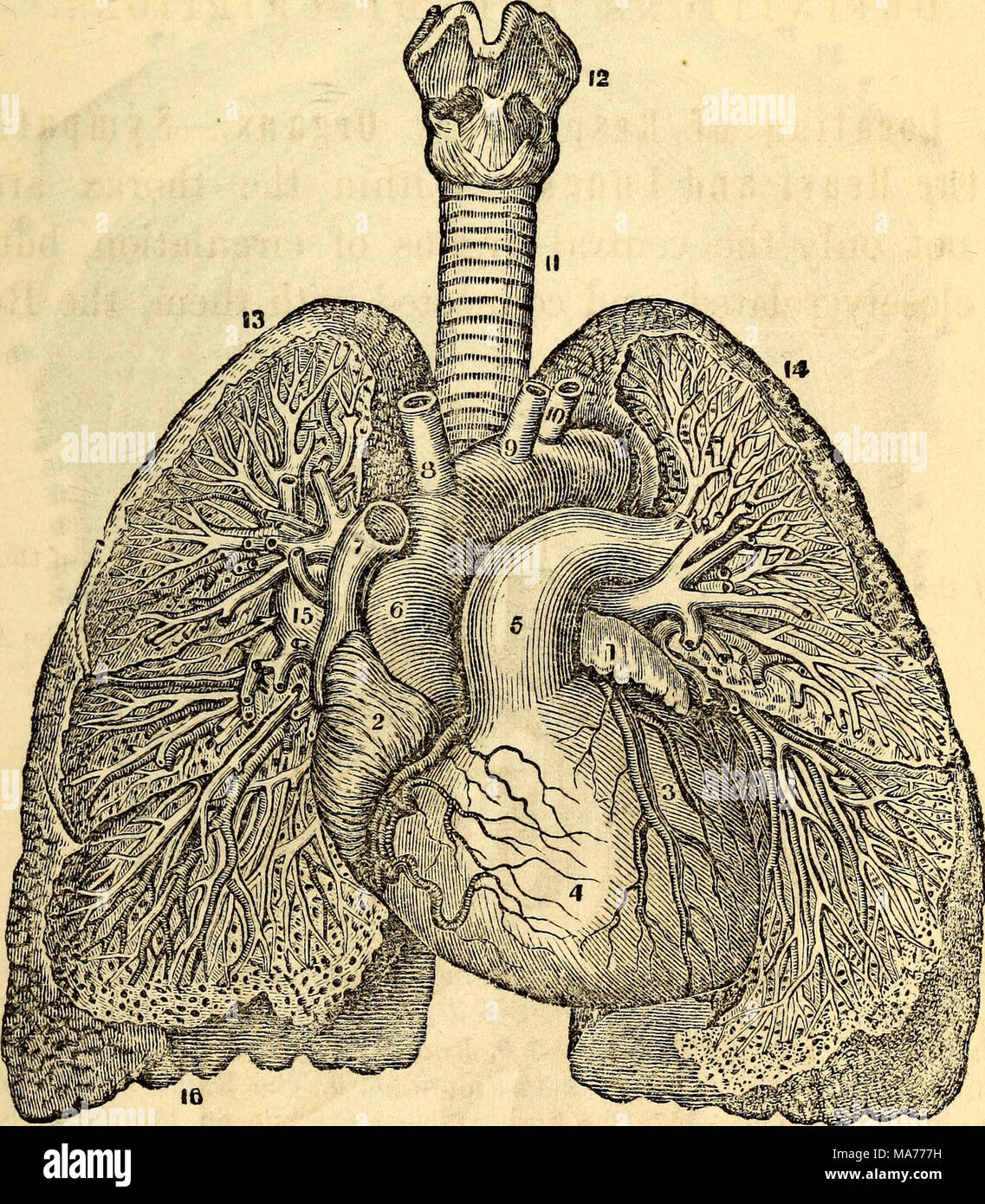
Vasos pulmonares.
La arteria pulmonar se extiende solo 5 cm más allá de la punta del ventrículo derecho y después se divide en las ramas principales derecha e izquierda, que vascularizan los dos pulmones correspondientes.
La arteria pulmonar tiene un grosor de pared un tercio del de la aorta. Las ramas de las arterias pulmonares son cortas, y todas las arterias pulmonares, incluso las arterias más pequeñas y las arteriolas, tienen diámetros mayores que sus correspondientes arterias sistémicas. Este aspecto, combinado con el hecho de que los vasos son delgados y distensibles, da al árbol arterial pulmonar una gran distensibilidad, que es en promedio de casi 7 ml/mmHg, que es similar a la de todo el árbol arterial sistémico. Esta gran distensibilidad permite que las arterias pulmonares se acomoden al gasto del volumen sistólico del ventrículo derecho.
Las venas pulmonares, al igual que las arterias pulmonares, también son cortas. Drenan inmediatamente la sangre que les llega hacia la aurícula izquierda.
Vasos bronquiales
La sangre también fluye hacia los pulmones a través de arterias bronquiales pequeñas que se originan en la circulación sistémica y transportan el 1-2% del gasto cardíaco total. Esta sangre arterial bronquial es sangre oxigenada, al contrario de la sangre parcialmente desoxigenada de las arterias pulmonares. Vascularizan los tejidos de soporte de los pulmones, como el tejido conjuntivo, los tabiques y los bronquios grandes y pequeños. Después de que esta sangre bronquial y arterial pase a través de los tejidos de soporte, drena hacia las venas pulmonares y entra en la aurícula izquierda, en lugar de regresar hacia la aurícula derecha. Por tanto, el flujo hacia la aurícula izquierda y el gasto del ventrículo izquierdo son aproximadamente un 1-2% mayores que el gasto del ventrículo derecho.
Linfáticos
Hay vasos linfáticos en todos los tejidos de soporte del pulmón, comenzando en los espacios tisulares conjuntivos que rodean a los bronquíolos terminales, y siguiendo hacia el hilio del pulmón, y desde aquí principalmente hacia el conducto linfático torácico derecho. Las sustancias en forma de partículas que entran en los alvéolos son retiradas parcialmente por medio de estos conductos, y también eliminan de los tejidos pulmonares las proteínas plasmáticas que escapan de los capilares pulmonares, contribuyendo de esta manera a prevenir el edema pulmonar.

PRESIONES EN EL SISTEMA PULMONAR
Curva de pulso de presión en el ventrículo derecho
La presión sistólica del ventrículo derecho del ser humano normal es en promedio de aproximadamente 25 mmHg, y la presión diastólica es en promedio de aproximadamente 0 a 1 mmHg, valores que son solo un quinto de los del ventrículo izquierdo.
Presiones en la arteria pulmonar
Durante la sístole la presión en la arteria pulmonar es esencialmente igual a la presión que hay en el ventrículo derecho.
Sin embargo, después del cierre de la válvula pulmonar al final de la sístole, la presión ventricular cae súbitamente, mientras que la presión arterial pulmonar disminuye más lentamente a medida que la sangre fluye a través de los capilares de los pulmones.
La presión arterial pulmonar sistólica se sitúa normalmente en promedio en unos 25 mmHg en el ser humano, la presión arterial pulmonar diastólica es de aproximadamente 8 mmHg y la presión arterial pulmonar media es de 15 mmHg.
Presión capilar pulmonar
La presión capilar pulmonar media, es de aproximadamente 7 mmHg. La importancia de esta baja presión capilar se analiza con más detalle más adelante en relación con las funciones de intercambio de líquidos de los capilares pulmonares.
Presiones auricular izquierda y venosa pulmonar

La presión media en la aurícula izquierda y en las venas pulmonares principales es en promedio de aproximadamente 2 mmHg en el ser humano en decúbito, y varía desde un valor tan bajo como 1 mmHg hasta uno tan elevado como 5 mmHg. Habitualmente no es posible medir la presión auricular izquierda de un ser humano utilizando un dispositivo de medida directa porque es difícil introducir un catéter a través de las cavidades cardíacas hacia la aurícula izquierda. Sin embargo, con frecuencia se puede estimar la presión auricular izquierda con una exactitud moderada midiendo la denominada presión de enclavamiento pulmonar. Esta medida se consigue introduciendo un catéter en
primer lugar a través de una vena periférica hasta la aurícula derecha, después a través del lado derecho del corazón y a través de la arteria pulmonar hacia una de las pequeñas ramas de la arteria pulmonar, y finalmente empujando el catéter hasta que se enclava firmemente en la rama pequeña.
La presión que se mide a través del catéter, denominada «presión de enclavamiento», es de aproximadamente 5 mmHg. Como todo el flujo sanguíneo se ha interrumpido en la arteria pequeña enclavada, y como los vasos sanguíneos que se extienden más allá de esta arteria establecen una conexión directa con los capilares pulmonares, esta presión de enclavamiento es habitualmente solo de 2 a 3 mmHg mayor que la presión auricular izquierda. Cuando la presión auricular izquierda aumenta a valores elevados, también lo hace la presión de enclavamiento pulmonar. Por tanto, las mediciones de la presión de enclavamiento se pueden utilizar para estudiar las alteraciones de la
presión capilar pulmonar y de la presión auricular izquierda en pacientes que tienen insuficiencia cardíaca congestiva.
VOLUMEN SANGUINEO EN PULMONES
El volumen de la sangre de los pulmones es de aproximadamente 450 ml, aproximadamente el 9% del volumen de sangre total de todo el aparato circulatorio. Aproximadamente 70 ml de este volumen de sangre pulmonar están en los capilares pulmonares, y el resto se divide aproximadamente por igual entre las arterias y las venas pulmonares.
Los pulmones sirven como reservorio de sangre
En varias situaciones fisiológicas y patológicas la cantidad de sangre de los pulmones puede variar desde tan poco como la mitad del valor normal hasta el doble de lo normal. Por ejemplo, cuando una persona sopla aire con tanta intensidad que se genera una presión elevada en los pulmones (como cuando se toca una trompeta), se pueden expulsar hasta 250 ml de sangre desde el aparato circulatorio pulmonar hacia la circulación sistémica. Por otro lado, la pérdida de sangre desde la circulación sistémica por una hemorragia puede ser compensada parcialmente por el desplazamiento automático de sangre desde los pulmones hacia los vasos sistémicos.
FLUJO SANGUÍNEO A TRAVÉS DE LOS PULMONES Y SU DISTRIBUCIÓN

El flujo sanguíneo a través de los pulmones es esencialmente igual al gasto cardíaco. Por tanto, los factores que controlan el gasto cardíaco también controlan el flujo sanguíneo pulmonar. En la mayoría de las situaciones los
vasos pulmonares actúan como tubos distensibles que se dilatan al aumentar la presión y se estrechan al disminuir la presión. Para que se produzca una aireación adecuada de la sangre, esta debe distribuirse a los segmentos de los pulmones en los que los alvéolos estén mejor oxigenados.
La disminucion del oxigeno alveolar reduce el flujo sanguineo alveolar local y regula la distribución del flujo sanguineo pulmonar

Cuando la concentración de O2 en el aire de los alvéolos disminuye por debajo de lo normal (especialmente cuando disminuye por debajo del 70% de lo normal [es decir, por debajo de 73 mmHg de Po2]) los vasos sanguíneos adyacentes se constriñen, con un aumento de la resistencia vascular de más de cinco veces a concentraciones de O2 muy bajas. Este efecto es opuesto al efecto que se observa en los vasos sistémicos, que se dilatan en lugar de constreñirse en respuesta a concentraciones bajas de O2. Aunque los mecanismos que promueven la vasoconstricción pulmonar durante la hipoxia no se conocen en profundidad, la baja concentración de O2 puede estimular la liberación de sustancias vasoconstrictoras o reducir la liberación de un vasodilatador, como el óxido nítrico, del tejido pulmonar.
El aumento en la resistencia vascular pulmonar como consecuencia de una baja concentración de O2 tiene una función importante de distribución del flujo sanguíneo allí donde sea más eficaz. Es decir, si algunos alvéolos están mal ventilados y tienen una concentración baja de O2, los vasos locales se constriñen. Esta constricción hace que la sangre fluya a través de otras zonas de los pulmones que están mejor aireadas, proporcionando de esta manera un sistema de control automático para distribuir el flujo sanguíneo a las zonas pulmonares en proporción a sus presiones alveolares de oxígeno.
ZONA 1, 2 Y 3 DEL FLUJO SANGUÍNEO PULMONAR
Los capilares de las paredes alveolares están distendidos por la presión de la sangre que hay en su interior, pero simultáneamente están comprimidos por la presión del aire alveolar que está en su exterior. Por tanto, siempre que la presión del aire alveolar pulmonar sea mayor que la presión de la sangre capilar, los capilares se cierran y no hay flujo sanguíneo. En diferentes situaciones normales y patológicas se puede encontrar una cualquiera de tres posibles zonas (patrones) del flujo sanguíneo pulmonar, como se señala a continuación:
Zona 1: ausencia de flujo durante todas las porciones del ciclo cardíaco porque la presión capilar alveolar local en esa zona del pulmón nunca aumenta por encima de la presión del aire alveolar en} ninguna fase del ciclo cardíaco.
Zona 2: flujo sanguíneo intermitente, solo durante los picos de presión arterial pulmonar, porque la presión sistólica en ese momento es mayor que la presión del aire alveolar, pero la presión diastólica es menor que la presión del aire alveolar.
Zona 3: flujo de sangre continuo, porque la presión capilar alveolar es mayor que la presión del aire alveolar durante todo el ciclo cardíaco.
El ejercicio aumenta el flujo sanguíneo a través de todas las partes de los pulmones
el flujo sanguíneo de todas las partes del pulmón aumenta durante el ejercicio. Una razón importante del aumento en el flujo sanguíneo es que durante el ejercicio las presiones vasculares pulmonares aumentan lo suficiente como para convertir los vértices pulmonares desde un patrón de flujo de zona 2 a un patrón de flujo de zona 3.
![Estos divertidos GIFs de Totoro son lo que necesitas para inspirarte a hacer ejercicio [Video] [Video] | Totoro, Emoticonos animados, Anime w](https://i.pinimg.com/originals/8d/31/1f/8d311ff5859de02df1da913ca14ef915.gif)
INTERCAMBIO CAPILAR DE LIQUIDO EN LOS PULMONES Y DINÁMICA DEL LÍQUIDO INTERSTICIAL
La dinámica del intercambio de líquido a través de las membranas capilares pulmonares es cualitativamente la misma que en los tejidos periféricos. Sin embargo, cuantitativamente hay diferencias importantes, como se señala a continuación:
- La presión capilar pulmonar es baja, de aproximadamente 7 mmHg, en comparación con una presión capilar funcional mucho mayor en los tejidos periféricos, de aproximadamente 17 mmHg.
- La presión del líquido intersticial del pulmón es ligeramente más negativa que en el tejido subcutáneo periférico. (Esta presión se ha medido de dos formas: con una micropipeta insertada en el intersticio pulmonar, que da un valor de aproximadamente –5 mmHg, y midiendo la presión de absorción de líquido desde los alvéolos, que da un valor de aproximadamente –8 mmHg.)
- La presión coloidosmótica del líquido intersticial pulmonar es de aproximadamente 14 mmHg, en comparación con menos de la mitad de este valor en los tejidos periféricos.
- Las paredes alveolares son muy delgadas, y el epitelio alveolar que recubre las superficies alveolares es tan débil que se puede romper si la presión positiva en los espacios intersticiales es mayor que la presión del aire alveolar (>0 mmHg), lo que permite el paso de líquido desde los espacios intersticiales hacia los alvéolos.
EDEMA PULMONAR
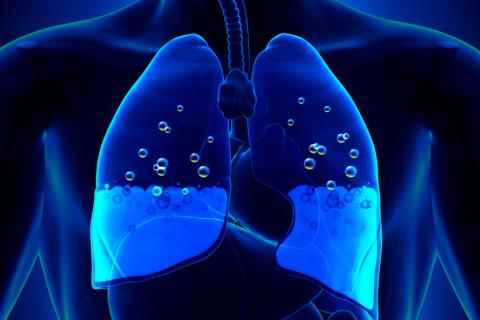
El edema pulmonar se produce de la misma forma en que se produce el edema en cualquier otra localización del cuerpo. Cualquier factor que aumente la filtración de líquido fuera de los capilares pulmonares o que impida la función linfática pulmonar y provoque un aumento de la presión del líquido intersticial pulmonar desde el intervalo negativo hasta el intervalo positivo dará lugar al
llenado rápido de los espacios intersticiales pulmonares y de los alvéolos con grandes cantidades de líquido libre.
Las causas más frecuentes de edema pulmonar son:
- Insuficiencia cardíaca izquierda o valvulopatía mitral, con los consiguientes grandes aumentos de la presión venosa pulmonar y de la presión capilar pulmonar y el encharcamiento de los espacios intersticiales y de los alvéolos.
- La lesión de las membranas de los capilares sanguíneos pulmonares producida por infecciones como la neumonía o por la inhalación de sustancias tóxicas como el gas cloro o el gas dióxido de azufre. Cada uno de estos mecanismos da lugar a una fuga rápida tanto de proteínas plasmáticas como de líquido desde los capilares hacia los espacios intersticiales pulmonares y los alvéolos.