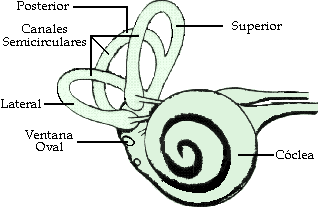
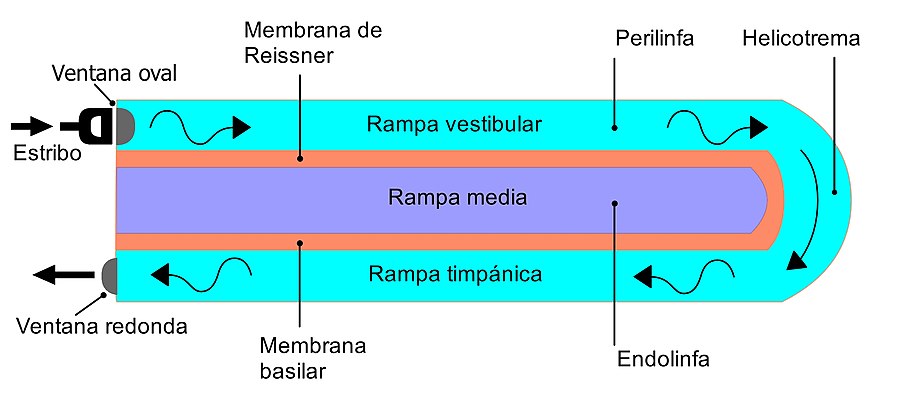
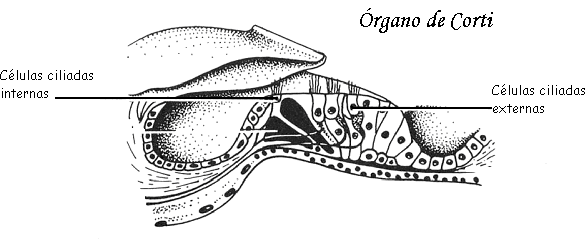
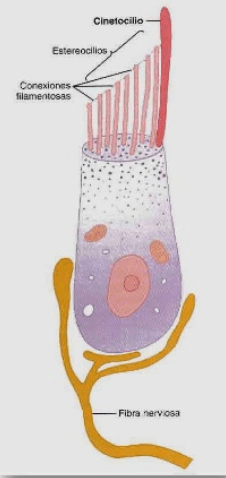
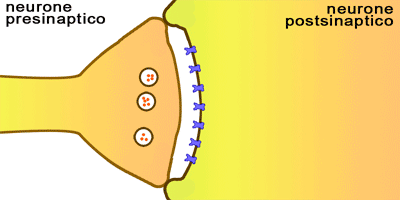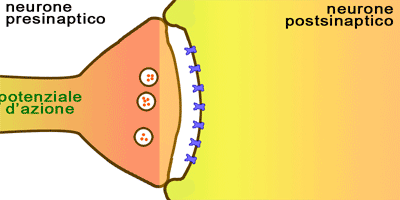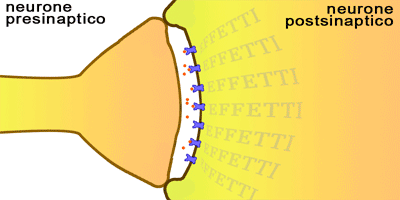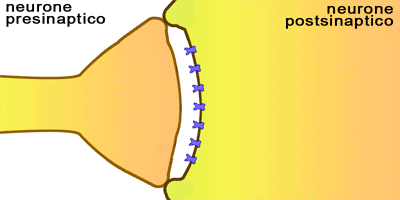
Ambos sentidos permiten distinguir los alimentos indeseables y los que son mortales de otros placenteros y nutritivos.
El sentido del gusto depende de las papilas gustativas. El olfato contribuye a la percepcion del gusto.

SENSACIONES GUSTATIVAS PRIMARIAS
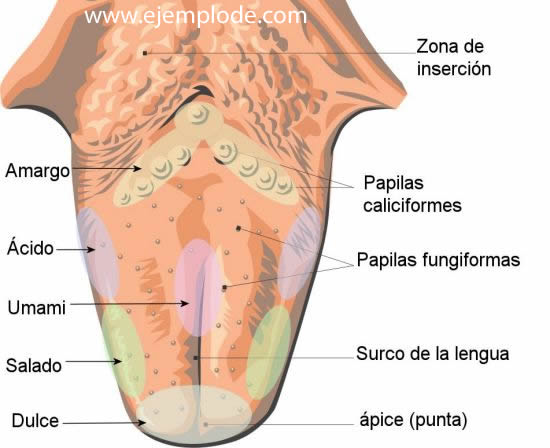
Se han agrupado cinco categorias denominadas «sensaciones gustativas primarias»
- Sabor agrio: se debe a las sustancias acidas y la intensidad del gusto es proporcional al logaritmo de la concentración de los iones de hidrogeno.
- Sabor salado: se atribuye, principalmente, a los cationes de la sales ionizadas, aunque las sales tambien activan otros receptores, lo que explica la ligera diferencia entre los alimentos de sabor salado.
- Sabor dulce: es el resultado de la activacion de diversos tipos de receptores, para azucares glicoles, alcoholoes, aldheídos y otros compuestos organicos.
- Sabor amargo: se debe a la activacion de varios receptores asociados a compouestos quimicos orgánicos. Las sustancias mas habituales son las sustancias hidrogenadas de cadena larga y alcaloides. Este grupo abarca sustancias medicinales como la quinina, la cafeina, la estrictina y la nicotin. El sabor fuerte se produce por un rechazo, qeu se debe al hehco de que las toxinas peligrosas de ciertos vegetales sean alcaloides.
- Sabor umami: la palabra «umami» proviene del japones, que significa delicioso, constituye la quinta categoria y el sabor dominante en todos los alientos que contienen L- glutamato, como los extractos cárnicos y el queso curado.

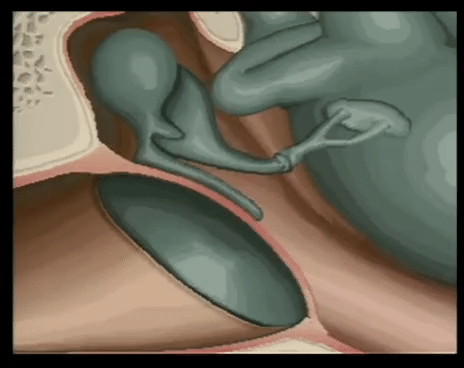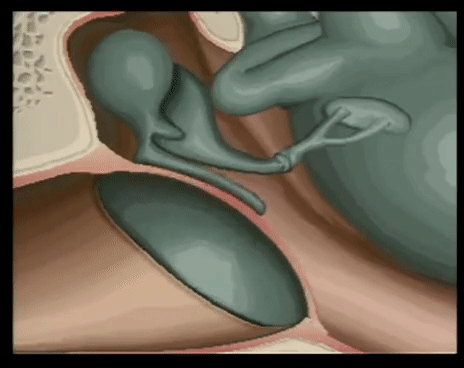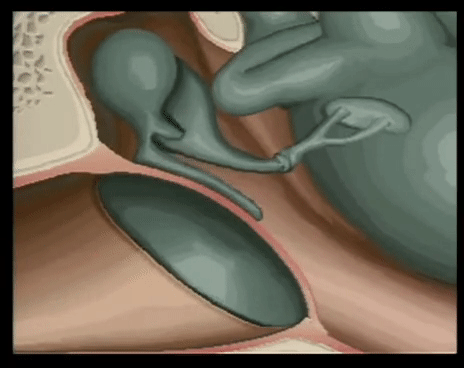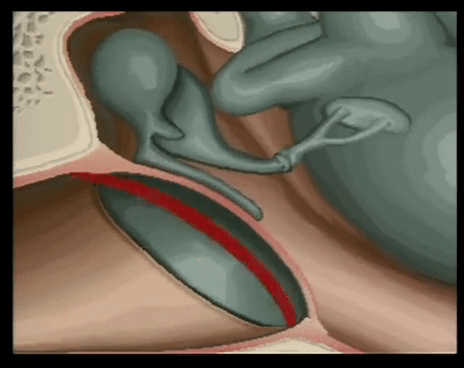
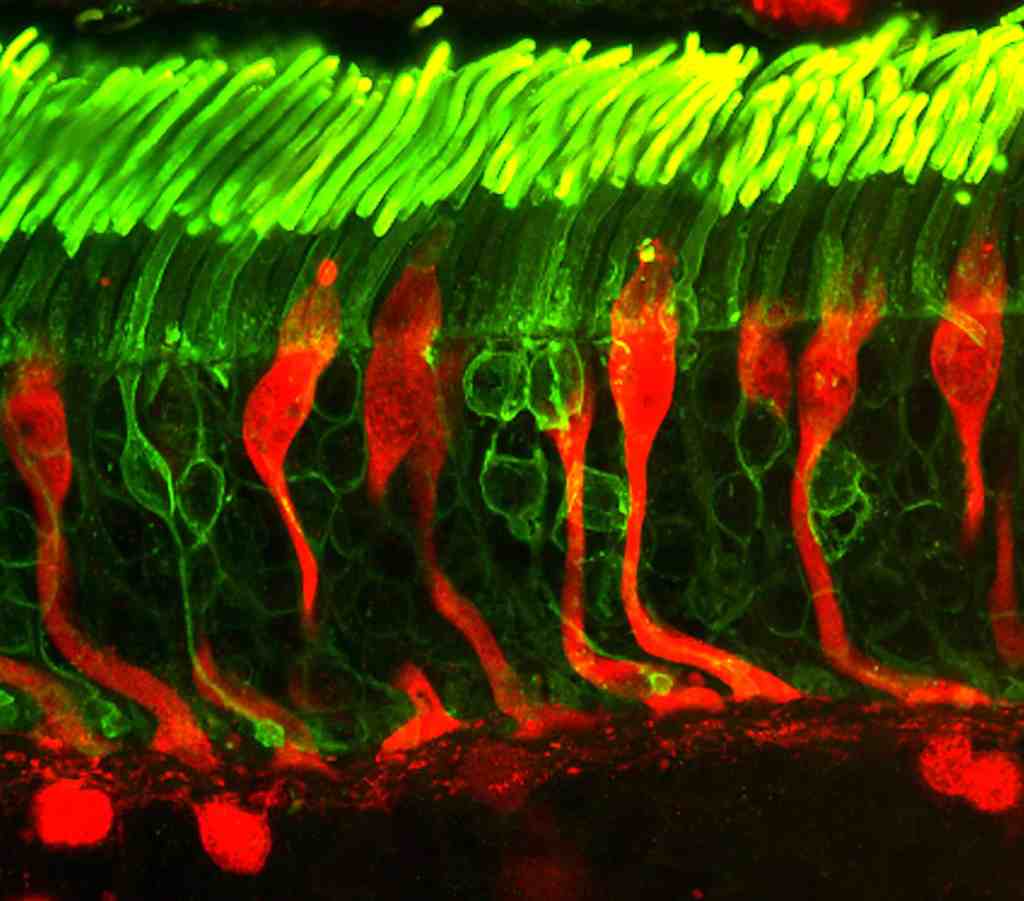
YEMAS GUSTATIVAS
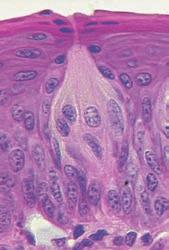
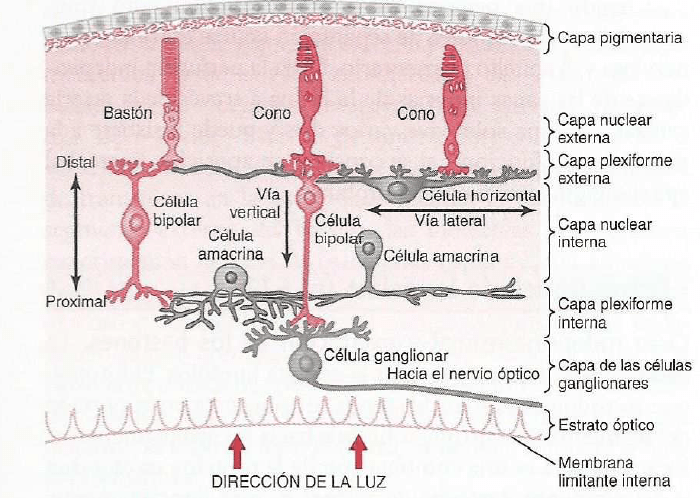
Se compones de unas 50 celulas epiteliales modificadas, algunas de las cuales, las celuals de sostén, cumplen con una función de soporte, mientras que otras son células receptoras verdaderas.
Las superficies apicales de las celulas gustativas se disponen alrededor de un poro gustativo.
Las microvellosidades o cilios gustativos sobresalesn del poro y constituyen la superficie receptora para las moleculas gustativas.
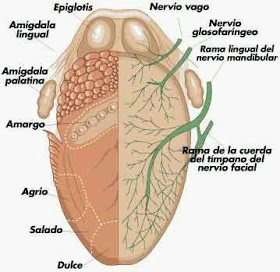
Las papilas fungiformes se encuentran en los dos tercios anteriores de la lengua.
Las papilas caliciformes forman un V en el tercio posterior de la lengua.
Laspapilas foliáceas se situan en los bordes laterales.
Cada yema gustativa responde de forma caracteristica a uo de los cinco sabores primarios.
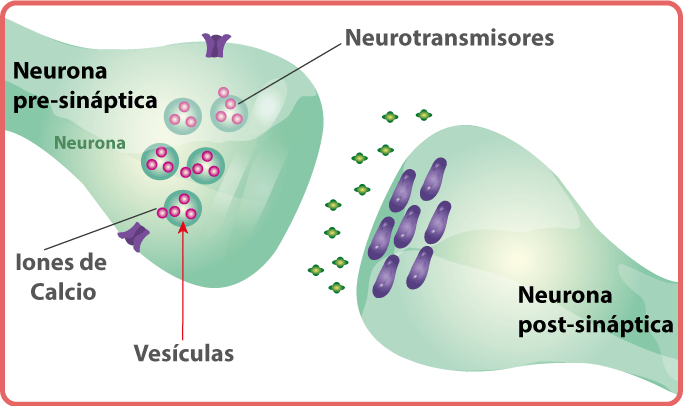
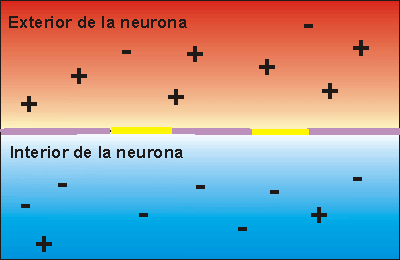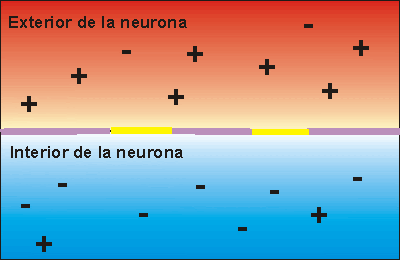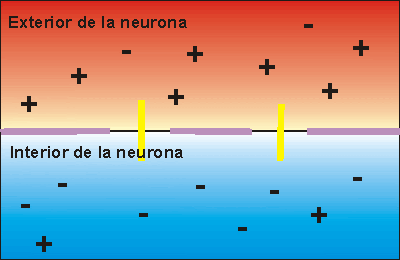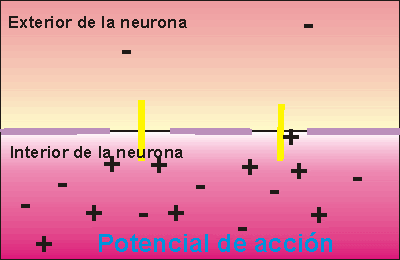
Las celulas receptoras, como otras celulas, tambien producen potencial un potencial de receptor. Cuando la sustancia entra a la papila, esta se despolariza, el grado de despolarizacion depende de la concetrancion de la sustancia que vaya a estimularla.
La union de la sustancia al receptor abre los canales ionicos especificos que permiten la entrada de sodio a la celula. Despues la sustancia gustativa es eliminada por la saliva.
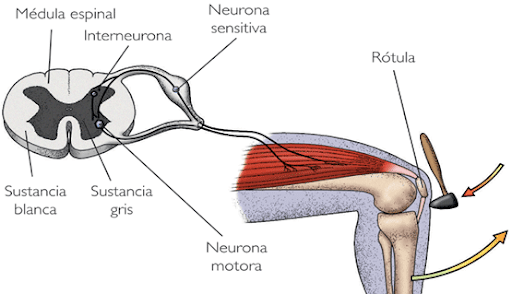
Las fibras gustativas de los dos tercios anteriores de la lengua viajan primero por las ramas del nervio trigemino y se unen despues a la cuerda del timpano, rama del nervio facial.
Las sensaciones gustativas del tercio posterior de la lengua son canalizadas por fibras del nervio glosofaringeo
Las fibras gustativas de la epiglotis dicurren por ramas del nervio vago.
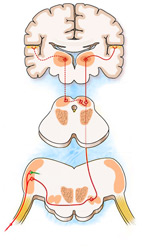
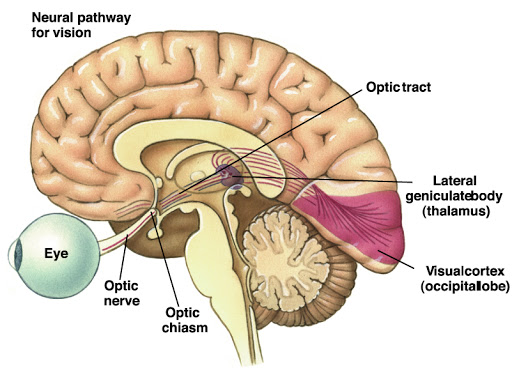
Todas las papilas pasan por el tracto solitario y acaban haciendo sinapsis en la parte ventral del nucleo del tracto solitario. Desde aqui los axones discurren en sentido rostral hasta alcanzar el nucleo ventromedial del tálamo hasta alcanzar la corteza cerebral, en concreto de la región ventral de la circunvoluncion poscentral, que se gira hasta la cisura de Silvio.
SENTIDO DEL OLFATO

El peor sentido del olfato conocido es el del humano, ya que hay especies animales en donde el olfato está mas desarrollado que en los humanos.
MEMBRANA OLFATORIA
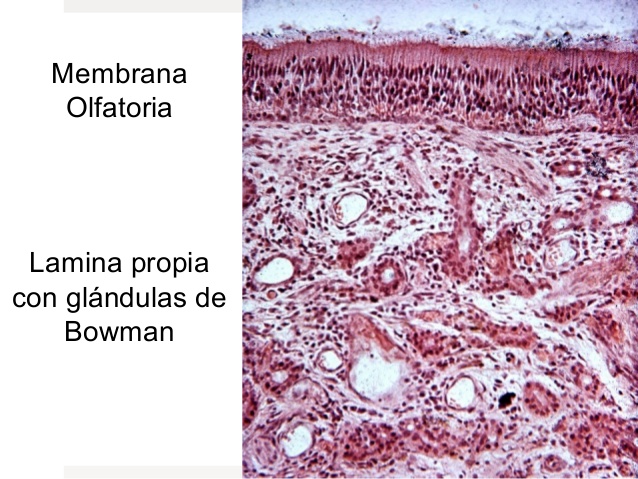
Se encuentra en la cavidad nasal y habitualmente se extiende solo 2,4 cm al cuadrado.
Las celulas olfatorias son neuronas bipolares del sistema nervioso central. Cada persona cuenta con alrededor de 100 millones de esas células, entremezcladas con un numero mucho menor de celulas se sostén.
La superficie apival de cada celula muestra alrededor de 4 a 25 cilios olfatorios, que contienen los receptores, y se proyecta al moco presente en la superficie epitelial.
Las grandulas de Bowman, que segregan moco a la superficie epitelial, se espacian por las celulas receptoras.
Para su estimulacion, las moleculas olorosas se difunden en el moco y se adhieren a la superficie receptora unidas a la proteina G citoplásmica.
se activa y se desprende la subunidad alpha de la proteina G, luego se activa en AMPc y se activan los canales de sodio en la celula, entonces entra el sodio en la celula, lo que provoca su despolarización.
La intensidad del estimulo olfatorio es proporcional al logaritmo de la concentracion del estimulo.
Se han descrito hasta 100 sensasiones olorosas, reducidas hasta siete olores primarios: alcanforado, almizcleño, floral, mentolado, etéreo, acre y pútrido.

TRANSMICION DE LAS SEÑALES OLFATORIAS EN EL SISTEMA NERVIOSO CENTRAL
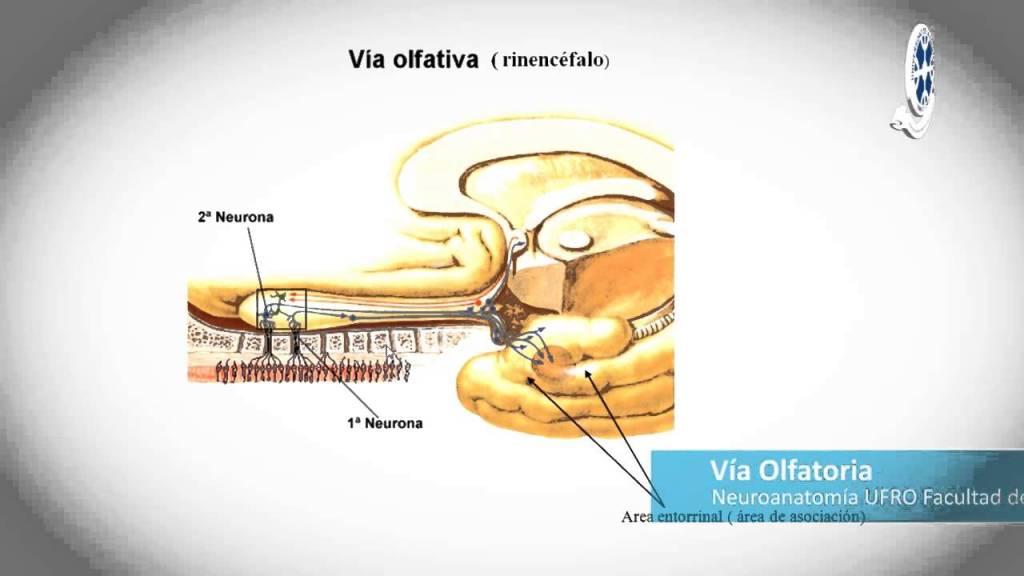
El bulbo olfatorio se situa sobre la lámina cribosa del etmoides, que separa las cavidades craneal y nasal.
Los nervios olfatorios que atraviesan las perforaciones de la lamina cribiforme y entran en el bulbo olfatorio, en los glomerulos, es decir, en ovillos de las dendritas de celulas mitrales y en penacho y fibras nerviosas olfatorias.
Los axones de las celulas mitrales y en penacho salen del bulbo olfatorio por el tracto olfatorio y entran en regiones especializadas de la corteza, sin hacer relevo al tálamo.
El AREA OLFATIVA MEDIAL está representada por los nucleos septales, que se proyectan al hipotalamo y otras regiones que regulan la conducta,. Se cree que este sistema interviene en funciones primitivas, como comerse los labios, salivar y otros comportamientos alimentarios.
El AREA OLFATORIA LATERAL se compone de las regiones prepiriforme, piriforme y cortical de los nucleos amigdalinos. Desde aqui, las señales se dirigen a las estructuras limbicas menos primitivas, como el hipocampo. Al parecer este sistema que asocia determinados olores con ciertas respuestas conductuales.
Otra via de filogenia mas reciente se proyecta aL nucleo dorsomedial del tálamo y luego a la corteza orbitofrontal.
Las fibras que se originan en el cerebro discurren de forma centrifuga hasta las celulas granulares del bulbo olfatorio. Estas ultimas inhiben las neuronas mitrales y en penacho del bulbo, con lo que se agudiza la capacidad para distinguir los diferentes olores.